|
Гормоны. Общий механизм действия.Гормоны – это продукты внутренней секреции, которые вырабатываются специальными железами или отдельными клетками, выделяются в кровь и разносятся по всему организму в норме вызывая определенный биологический эффект. Сами гормоны непосредственно не влияют на какие-либо реакции клетки. Только связавшись с определенным, свойственным только ему рецептором вызывается определенная реакция. Гормоны имеют различную химическую структуру. Это приводит к тому, что они имеют разные физические свойства. Гормоны разделяют на водо- и жирорастворимые. Принадлежность к какому-то из этих классов обуславливает их механизм действия. Это объясняется тем, что жирорастворимые гормоны могут спокойно проникать через клеточную мембрану, которая состоит преимущественно из бислоя липидов, а водорастворимые этого не могут. В связи с этим рецепторы(Р) для водо- и жирорастворимых гормонов имеют различное место локализации (мембрана и цитоплазма). Связавшись с мембранным рецептором гормон вызывает каскад реакций в самой клетке, но никак не влияет на генетический материал. Комплекс цитоплазматического Р и гормона может воздействовать на ядерные рецепторы и вызывать изменения в генетическом аппарате, что ведет к синтезу новых белков. Рассмотрим это поподробнее. Механизм действия стероидных (жирорастворимых) гормонов
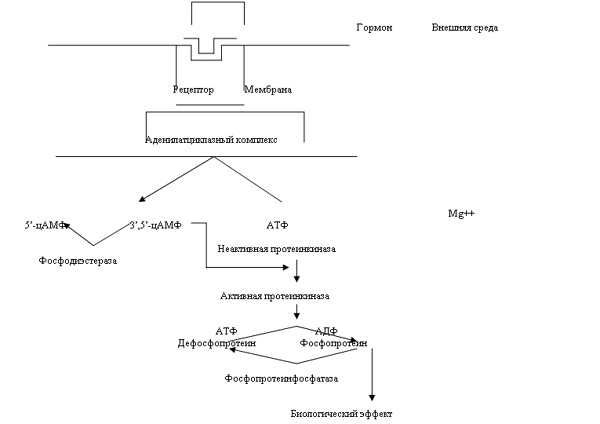
Вторичные посредники Пептидные гормоны, амины и нейромедиаторы в отличие от стероидов ¾ гидрофильные соединения и не способны легко проникать через плазматическую мембрану клетки. Поэтому они взаимодействуют с расположенными на поверхности клетки мембранными рецепторами. Гормон-рецепторное взаимодействие иницирует высококоординированную биологическую реакцию, в которой могут участвовать многие клеточные компоненты, причем некоторые из них расположены на значительном расстоянии от плазматической мембраны. цАМФ ¾ первое соединение, которое открывший его Сазерленд назвал «вторым посредником», потому что «первым посредником» он считал сам гормон, вызывающий внутриклеточный синтез «второго посредника», который опосредует биологический эффект первого. На сегодняшний день можно назвать не менее 3 типов вторичных посредников: 1)циклические нуклеотиды (цАМФ и цГМФ); 2)ионы Ca и 3)метаболиты фосфатидилинозитола. С помощью таких систем небольшое число молекул гормона, связываясь с рецепторами, вызывает продукцию гораздо большего числа молекул второго посредника, а последние в свою очередь влияют на активность еще большего числа белковых молекул. Таким образом, происходит прогрессивная амплификация сигнала, исхдно возникающего при связывании гормона с рецептором. Упрощенно действие гормона через цАМФ можно представить так:
 1.Следует отметить, что рецепторы тоже являются динамическими структурами. Это означает, что их количество может или снижаться или повышаться. Например у людей с увеличенной массой тела уменьшается количество рецепторов инсулина. Опыты показали, что при нормализации их массы замечается увеличение количества рецепторов до нормального уровня. Иными словами, при повышении или снижении концентрации инсулина имеют место реципрокные изменения концентрации рецепторов. Считается, что это явление может защитить клетку от слишком интенсивной стимуляции при неадекватно высоком уровне гормона.
2.Активация аденилатциклазы (А) тоже является регулируемым процессом. Ранее считалось, что гормон (Г), связываясь с рецептором (Р), изменяет его конформацию, что приводит к активированию А. Однако оказалось, что А, является аллостерическим ферментом, который активируется под действием ГТФ. ГТФ переносит специальный белок (трансдуктор) G. В связи с этим была принята модель , описывающая не только активацию А, но и терминацию этого процесса
Таким образом, «выключающим» систему сигналом служит гидролиз ГТФ. Для возобновления цикла ГДФ должен отсоединиться от G, что происходит при связывании гормона с Р.
Некоторые факторы оказывают ингибирующее действие на А и вызывают снижение концентрации цАМФ. Примерами агонистов стимулирующих циклазу, могут служить глюкагон, АДГ, ЛГ, ФСГ, ТТГ и АКТГ. К факторам ингибирующим циклазу, относятся опиоиды, соматостатин, ангиотензин II и ацетилхолин. Адреналин может как стимулировать (через b-рецепторы), так и ингибировать (через a-рецепторы) данный фермент. Возникает вопрос каким же образом осуществляется двунаправленная регуляция А. Оказалось, что ингибирующая система включает в себя трехмерный белок, чрезвычайно похожий на приведенный выше G-белок. Эффект Gи может быть описан следующим образом:
После фосфорилирования белков-ферментов в ходе выше описанных реакций (см. рис 1) изменяется их конформация. Следовательно изменяется и конформация их активного центра, что ведет к их активированию или ингибированию. Получается, что благодаря вторичному посреднику цАМФ в клетке активируются или ингибируется действие специфичных для нее ферментов, что вызывает определенный биологический эффект свойственный для этой клетки. В связи с этим несмотря на большое количество ферментов, которые действуют через вторичный посредник цАМФ, в клетке возникает определенный, специфический ответ.
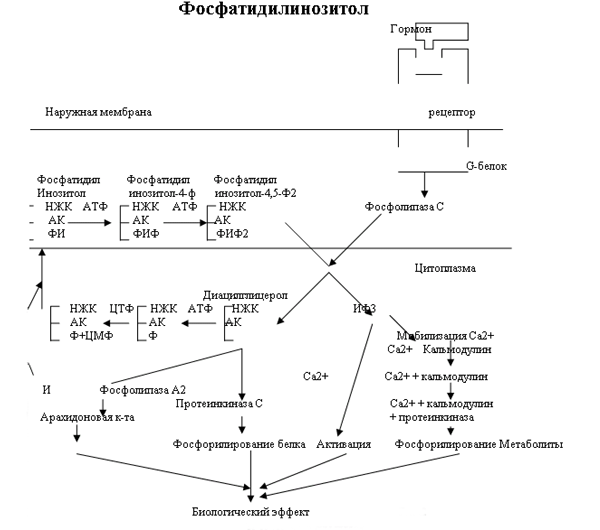
КальмодулинЭффект кальмодулина очень похож на эффект цАМФ. В клетке по мере дифференцировки образуется набор Са2+-кальмодулин-чувствительных белков. Кальмодулин активирует ферменты связываясь с ними и изменяя их конформацию. В то же время кальмодулин сам является аллостерическим белком, который проявляет свои биологические свойства только в связанном с ионами Са виде. Комплекс кальмодулина с Са может изменять активность белков одним из двух способов:
1) путем прямого воздействия на фермент-мишень
Взаимодействие вторых посредников Как было показано на рис.2 Фосфатидилинозитол также запускает кальмодулин. В то же время кальмодулин как и цАМФ активируют белки-ферменты благодаря чему вызывается биологический эффект. Даже среди 3 известных на сегодняшний день типов вторичных посредников мы видим тесное взаимодействие. Рассмотрим взаимодействие кальмодулина и цАМФ. Большое разнообразие набора кальмодулиновых белков в клетке ведет к различному действию на цАМФ в разных клетках организма. В одном случае кальмодулин активирует фосфодиэстеразу (см. рис.1) что приводит к ингибированию эффекта цАМФ. В другом случае он активирует аденилатциклазу, в третьем ингибирует. В связи с таким разнообразием эффектов Расмуссен предположил 5 различных ситуаций, в которых конечная интегральная реакция зависит от одновременного присутствия цАМФ и Са2+, действующих вместе или последовательно:
Гормон роста (соматотропин)Как и каждый гормон аденогипофиза соматотропин синтезируется под действием рилизинг фактора (ГР-РФ). Стимуляторами выработки ГР-РФ являются тиреоидный гормоны и глюкокортикоиды. Главным ингибитором реактивности питуицитов по отношению к ГР-РФ является соматостатин. Он стимулируется соматомединами. То есть можно сказать что соматостатин действует по методу обратной связи. ГР-РГ стимулирует как секрецию так и синтез СТГ. Этот эффект опосредуется связыванием ГР-РФ с рецепторами плазматической мембраны соматотрофов и активацией трех систем вторичных посредников. С их помощью происходит усиленный синтез мРНК СТГ. Этот эффект действия водорастворимого гормона на ядерный аппарат клетки объясняют фосфорилированием и дефосфорилированием определенной группы протеинкиназ, которые действуют на генетический аппарат клетки. На данный момент ни одна из этих протеинкиназ не идентифицирована. СТГ не действует прямо на клетку. Он переносится кровью в печень где превращается в соматомедины. То что СТГ не действует прямо на гормон было доказано на опыте, когда изолированную хрящевую ткань перфузировали раствором, который содержал большое количество СТГ и не наблюдали никакого ее роста. При помещении ее в нормальную сыворотку крови ее клетки росли. Потом были найдены соматомедины, которые непосредственно вызывают эффект роста. Клеточный механизм действия СТГ на хрящевую ткань связан с эффектами соматомедина. Хотя стимуляция роста и других тканей, таких как печень и мышцы, может опосредоваться аналогичным механизмом, in vitro показано и прямое действие СТГ на мышцу. Так, подобно инсулину СТГ вызывает независимое от синтеза белка усиление транспорта глюкозы. Спустя некоторое время после добавления СТГ мышца приобретает резистентность к действию инсулина, и в этот период усиливается транспорт аминокислот, зависящий от синтеза нового белка. Действие СТГ на печень до сих пор недостаточно изучено. Гормон стимулирует синтез белка, РНК. Это в свою очередь ведет к увеличению количества полисом. Также в конце концов он стимулирует синтез ДНК. На данный момент неизвестно, как СТГ, имеющий рецепторы на мембране может вызывать эффекты похожие на действие стероидных гормонов. Можно предположить, что он вызывает такие эффекты вызывая синтез ферментов, отвечающих за синтез нуклеиновых кислот. Это также доказывает то, что жирорастворимые гормоны включают только определенные гены, а СТГ ведет к генерализованой реакции увеличения синтеза белка на включенных генах, что ведет к росту клетки, а не к ее дифференциации. До сих пор не вполне ясно, какие эффекты вызывает непосредственно СТГ, а какие соматомедины. Известно точно, что соматомедины действуют непосредственно на хрящевую ткань, вызывая в хондробластах:
Тиреоидные гормоны (Т3 – Т4)Пептидергические нейроны в преоптической области гипоталамуса синтезируют и выделяют в воротную систему гипофиза тиреотропин-рилизинг гормон (ТРГ). ТРГ стимулирует выработку в клетках гипофиза тиреотропного гормона (ТТГ). ТТГ представляет собой гликопротеид с молекулярной массой 29000. ТТГ может влиять на развитие тиреоидных фолликулярных клеток, предшествующих образованию самих фолликулов в щитовидной железе. Так, было показано, что тиреоидные клетки in vitro, диспергированные с помощью протеолитических ферментов, при добавлении в среду ТТГ вновь собираются в молекулы. Это показывает, что ТТГ способствует синтезу каких-то компонентов клеточной мембраны, необходимых для распознавания друг друга. ТТГ-рецепторный комплекс, образующийся в результате связывания ТТГ, вызывает ответные биологические реакции с помощью не менее четырех внутриклеточных посредников: цАМФ, инозитолтрифосфата, диацилглицерола и комплекса Са2+-кальмодулин. Интересно, что активация аденилатциклазы в мембранах тиреоидных клеток под действием ТТГ в основном обуславливается ингибированием Gи-белка. Под влиянием ТТГ в клетках тиреоидных фолликулов возникают глубокие морфологические изменения. Тиреоидная клетка в высокой степени полярна: на ее апикальной поверхности имеются многочисленные микроворсинки (увеличение площади), к которым прилегают запасы коллоида в фолликуле. Базальная поверхность клетки контактирует с кровью. Коллоид представляет собой белковый раствор богатый на остаток аминокислоты тирозина. В ходе биохимических реакций фенольный гидроксид тирозина соединяется с фенолом другого тирозина и присоединяя к себе йод образуется тетрайодтиронин, связанный пептидными связями с остатками других аминокислот. Такие белки называются тиреоглобулинами.
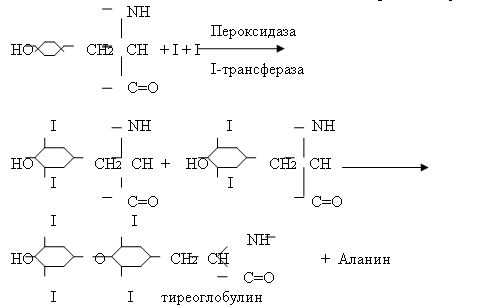 Под действием ТТГ в клетку поступают ионы Са, которые связываясь с кальмодулином вызывают движение лизосом из базальной части клетки к апикальной. Здесь происходит слияние лизосомы с каплей коллоидного раствора и образующаяся в результате фаголизосома движется в обратном направлении. При этом имеющийся в ней набор гидролаз разщепляет пептидные связи и образуются тироксин и трийодтиронин, которые и попадают в кровь. Второй стадией действия ТТГ-рецепторного комплекса является выработка тиреоглобулина. В данном случае эффект ТТГ на фолликулярную клетку похож на действие СТГ. В ходе усиления синтеза РНК, повышения проникновения глюкозы в клетку синтезируется I-содержащий белок тиреоглобулин.
Тиреоидные гормоны в кровиПосле того как протеаза тиреоглобулина высвобождает занесенные в фолликулярном коллоиде T4 и Т3 они выходят из клетки и попадают в кровоток, где избирательно связываются с одним из нескольких белков-переносчиков. Считается, что при попадении в клетку тиреоидные гормоны диссоциируют с белком переносчиком. Биологическое действие тиреоидных гормонов Недостаточность и избыток тиреоидных гормонов сказывается практически на любом процессе жизнедеятельности. В целом, чем раньше возникает дефицит гормона, тем сильнее это сказывается на ЦНС. Если такой дефицит долгое время остается без изменений, то могут наступить необратимые нарушения ЦНС. Даже возникновение гормональной недостаточности у ранее здорового взрослого человека ведет к появлению тех же симптомов психической иннертности, апатии и сонливости, которые характерны для ребенка с кретинизмом. Больной тиреотоксикозом ¾ возбужденный, беспокойный, эмоционально лабильный. Больной гипотиреозом предпочитает теплую погоду и сильно страдает от холода; при гипертиреозе ситуация обратная. Он «чрезмерно» приспособлен к холоду и плохо переносит тепло. По существу в тепле у него не выключается механизм адаптации к холоду. Нет такой системы органов, на которую не влиял бы дефицит или избыток тиреоидных гормонов. Поведенческие сдвиги, вызванные патологией щитовидной железы, отражают действие гормонов на ЦНС и нейромышечный аппарат. При гипертиреозе наблюдается мышечная слабость, усиленная всасываемость глюкозы, повышение ХОК, увеличение основного объема. При гипотиреозе наоборот. Клеточные механизмы действия тиреоидных гормонов Механизм действия тиреоидных гормонов не укладывается в обычные схемы, разработанные для стероидных гормонов, аминов и пептидных гормонов. От аминов и пептидов тиреоидные гормоны отличаются своей растворимостью в липидах, чем напоминают стероидные гормоны. Также необходимо проводить различие между ролью тиреоидных гормонов в процессе дифференцировки и их ролью в поддержании функций полностью дифференцированных клеток.
1. Периферическое дейодирование Т4
2. Действие на плазматическую мембрану
3. Действие на митохондрии
4. Ядро
С учетом того, что тиреоидные гормоны действуют на увеличение транспорта аминокислот, увеличение количества АТФ, то синтез новых белков происходит достаточно быстро. Избирательная стимуляция синтеза определенных иРНК ведет к дифференциации клетки в детстве, а в зрелом возрасте в поддержании ее нормального функционирования. Этот эффект очень сильно заметен по отношению к ЦНС, потому что нарушение образования тиреоидных гормонов в первую очередь стает заметным по изменениям поведения, психики и эмоциональности.
Вазопресин (АДГ)Антидиуретический гормон (АДГ) ¾ это только один из компонентов сложного комплекса нейрональных, эндокринных и поведенческих механизмов, совместное действие которых обеспечивает гомеостаз жидкости и электролитов в организме. Однако на первой линии обороны гомеостаза располагаются АДГ, ренин-ангиотензин-альдостерон. Между тремя механизмами поддержания постоянства жидкости, электролитов и объема (АДГ, ренин-ангиотензин-альдостероновая система, жажда и питьевое поведение) тесная связь.
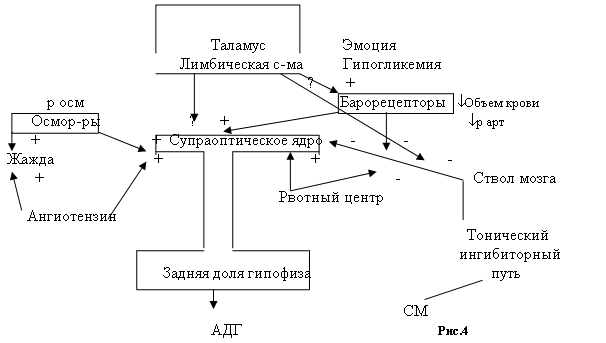
Клеточные механизмы действия АДГ
Рецепторы АДГ в почках известны как V2-рецепторы, а в кровеносных сосудах и печени как V1-рецепторы. АДГ активирует разные эффекторные системы и тем самым опосредуют разные биологические эффекты. V2-рецепторы чувствительных к АДГ клеток почечных канальцев расположены на контрлюминальной (обращенной к крови и лимфе) поверхности канальца. В клеточной мембране они взаимодействуют с комплексом G-белок-аденилатциклаза и стимулируют образование цАМФ. Наиболее отчетливый биологический эффект повышенного уровня цАМФ регистрируется на люминальной мембране с противоположной стороны клетки. В отсутствии АДГ люминальные мембраны чувствительных клеток практически не проницаемы для воды. В результате образуется гипотоническая моча, т.е. утрачивается ион-концентрирующая способность почек. При взаимодействии АДГ с рецепторами увеличивается концентрация цАМФ, фосфорилируются какие-то невыявленные на данный момент белки, и отдельные белковые частицы перемещаются из глубины клетки к ее люминальной мембране, где собираются в агрегаты. Эти примечательные частицы придают ранее водонепроницаемой мембране способность транспортировать в клетку воду, свободную от ионов. Вода идет по градиенту концентраций в кровеносное русло. Биологический эффект в клетках почечных канальцев полноценно проходит в присутствии кальмодулина, так как для него надо перенос определенных белковых комплексов, который происходит при наличии кальмодулина, который обеспечивает транспорт частиц микротрубочками. Реакция сосудов заключается в сокращении их гладкомышечного слоя и должна поэтому опосредоваться увеличением концентрации Са2+ в цитозоле. В реакции участвует и кальмодулин.
Система ренин-ангиотензин-альдостеронЭта система тесно связана с юкстагломерулярным аппаратом почки. Главная функция юкстагломерулярного аппарата ¾ регуляция артериального давления и объема крови. Кроме того в этот аппарат поступает информация о концентрации Na+ в канальцевой моче; нервная регуляция осуществляется симпатическими нервами через b-рецепторы. В ответ на снижение давления, объема крови и концентрации натрия выделяется ренин. Он воздействует на a2-глобулины и расщепляет их с образованием ангиотензина-1. Затем превращающий фермент, который находится в легких, отщепляет от ангиотензина 1 две терминальные аминокислоты с образованием ангиотензина 2. Этот октапептид вызывает главным образом сужение артериол. Одновременно он выступает тропным гормоном для клеток клубочковой зоны, синтезирующих и секретирующих альдостерон. Секретирующие альдостерон клетки надпочечников находятся под контролем целого комплекса факторов, в том числе концентрации электролитов в среде и уровня АКТГ. Альдостерон способствует реабсорбции натрия из клубочкового фильтрата в почечных канальцах и увеличению экскреции калия с мочой. Задержка натрия имеет по меньшей мере два следствия: во-первых, увеличивается задержка воды и потому восстанавливается объем жидкости; во-вторых, повышенная концентрация натрия придает мышечным клеткам стенок артериол большую чувствительность к вазоактивным веществам. Эти эффекты дополняют друг друга и в конечном счете восстанавливают артериальное давление до стабильного уровня.
Стимуляторы синтеза альдостерона
Повышение концентрации К+ тоже стимулирует синтез и секрецию альдостерона; в основе лежит деполяризация мембраны клеток клубочковой зоны и соответственно открытие зависимых от вольтажа кальциевых каналов. Деполяризация активирует и аденилатциклазную систему, что приводит к умеренному повышению уровня цАМФ.
Механизм действия альдостерона
ГлюкокортикоидыГлюкокортикоиды крови транспортируются с помощью кортикостероид-связывающего глобулина. Кортизол – основной представитель так называемых глюкокортикоидов, т.е. стероидов надпочечников, действующих преимущественно на метаболизм органических соединений. Глюкокортикоиды прямо или опосредованно регулируют практически все физиологические и биохимические процессы. Кортизол способен изменять реактивность клеток по отношению к другим гормонам и нейромедиаторам. Для того чтоб понять биологическое действие глюкокортикоидов можно привести в пример восстановленные нарушения при введении заместительных доз глюкокортикоидов при дефиците кортизола:
В больших дозах глюкокортикоиды ингибируют почти все фазы воспалительного процесса. Они блокируют расширение капилляров, адгезию и миграцию лейкоцитов, секрецию гистамина и серотонина, образования кининов и т.д. С выяснением роли простагландинов была доказана антивоспалительная роль глюкокортикоидов. Они тормозят синтез простагландинов и лейкотриенов in vivo. Клеточный механизм действия (см. механизм действия стероидных гормонов). |