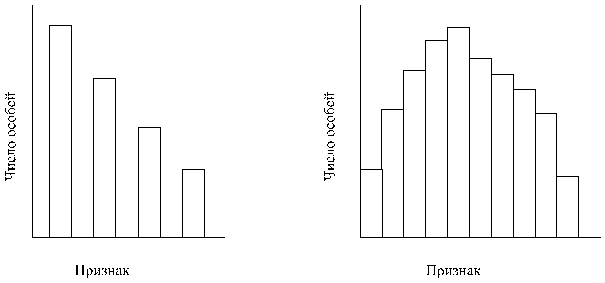|
Генетика
Генетика по праву может считаться одной из самых важных областей биологии. На протяжении тысячелетий человек пользовался генетическими методами для улучшения домашних животных и возделываемых растений, не имея представления о механизмах, лежащих в основе этих методов. Судя по разнообразным археологическим данным, уже 6000 лет назад люди понимали, что некоторые физические признаки могут передаваться от одного поколения другому. Отбирая определенные организмы из природных популяций и скрещивая их между собой, человек создавал улучшенные сорта растений и породы животных, обладавшие нужными ему свойствами. Однако лишь в начале XX в. ученые стали осознавать в полной мере важность законов наследственности и ее механизмов. Хотя успехи микроскопии позволили установить, что наследственные признаки передаются из поколения в поколение через сперматозоиды и яйцеклетки, оставалось неясным, каким образом мельчайшие частицы протоплазмы могут нести в себе «задатки» того огромного множества признаков, из которых слагается каждый отдельный организм. Первый действительно научный шаг вперед в изучении наследственности был сделан австрийским монахом Грегором Менделем, который в 1866 г. опубликовал статью, заложившую основы современной генетики. Мендель показал, что наследственные задатки не смешиваются, а передаются от родителей потомкам в виде дискретных (обособленных) единиц. Эти единицы, представленные у особей парами, остаются дискретными и передаются последующим поколениям в мужских и женских гаметах, каждая из которых содержит по одной единице из каждой пары. В 1909 г. датский ботаник Иогансен назвал эти единицы гедам», а в 1912 г. американский генетик Морган показал, что они находятся в хромосомах. С тех пор генетика достигла больших успехов в объяснении природы наследственности и на уровне организма, и на уровне гена.
Природа геновИзучение наследственности уже давно было связано с преставлением о ее корпускулярной природе. В 1866 г. Мендель высказал предположение, что признаки организмов определяются наследуемыми единицами, которые он назвал “элементами”. Позднее их стали называть “факторами” и, наконец, генами; было показано, что гены находятся в хромосомах, с которыми они и передаются от одного поколения к другому. Несмотря на то, что уже многое известно о хромосомах и структуре ДНК, дать определение гена очень трудно, пока удалось сформулировать только три возможных определения гена:
Исследования Менделя
Грегор Мендель родился в Моравии в 1822 г. В 1843 г. он поступил в монастырь августинцев в Брюние (ныне Брно, Чехословакия), где принял духовный сан. Позже он отправился в Вену, где провел два года, изучая в университете естественную историю и математику, после чего в 1853 г. вернулся в монастырь. Такой выбор предметов, несомненно, оказал существенное влияние на его последующие работы по наследованию признаков у гороха. Будучи в Вене, Мендель заинтересовался процессом гибридизации растений и, в частности, разными типами гибридных потомков и их статистическими соотношениями. Эти проблемы и явились предметом научных исследований Менделя, которые он начал летом 1856 г. Успехи, достигнутые Менделем, частично обусловлены удачным выбором объекта для экспериментов-гороха огородного (Рisum sativum). Мендель удостоверился, что по сравнению с другими этот вид обладает следующими преимуществами:
И до Менделя многие ученые проводили подобные эксперименты на растениях, но ни один из них не получил таких точных и подробных данных; кроме того, они не смогли объяснить свои результаты с точки зрения механизма наследственности. Моменты, обеспечившие Менделю успех, следует признать необходимыми условиями проведения всякого научного исследования и принять их в качестве образца. Условия эти можно сформулировать следующим образом:
Как писал Мендель, «достоверность и полезность всякого эксперимента определяются пригодностью данного материала для тех целей, в которых он используется». Следует, однако, отметить, что в выборе экспериментального объекта Менделю кое в чем и просто повезло: в наследовании отобранных им признаков не было ряда более сложных особенностей, открытых позднее, таких как неполное доминирование, зависимость более чем от одной пары генов, сцепление генов.
Наследование при моногибридном скрещивании и закон расщепленияДля своих первых экспериментов Мендель выбирал растения двух сортов, четко различавшихся по какому-либо признаку, например по расположению цветков: цветки могут быть распределены по всему стеблю (пазушные) или находиться на конце стебля (верхушечные). Растения, различающиеся по одной паре альтернативных признаков, Мендель выращивал на протяжении ряда поколений. Семена от пазушных цветков всегда давали растения с пазушными цветками, а семена от верхушечных цветков- растения с верхушечными цветками. Таким образом, Мендель убедился, что выбранные им растения размножаются в чистоте (т.е. без расщепления потомства) и пригодны для проведения опытов по гибридизации (экспериментальных скрещиваний). Его метод состоял в следующем: он удалял у ряда растений одного сорта пыльники до того, как могло произойти самоопыление (эти растения Мендель называл «женскими»); пользуясь кисточкой, он наносил на рыльца этих «женских» цветков пыльцу из пыльников растения другого сорта; затем он надевал на искусственно опыленные цветки маленькие колпачки, чтобы на их рыльца не могла попасть пыльца с других растений. Мендель проводил реципрокные скрещивания - переносил пыльцевые зерна как с пазушных цветков на верхушечные, так и с верхушечных на пазушные. Во всех случаях из семян, собранных от полученных гибридов, вырастали растения с пазушными цветками. Этот признак-«пазушные цветки»,-наблюдаемый у растений первого гибридного поколения, Мендель назвал доминантным; позднее, в 1902 г., Бэтсон и Сондерс стали обозначать первое поколение гибридного потомства символом F1. Ни у одного из растений F1 не было верхушечных цветков. На цветки растений F1 Мендель надел колпачки (чтобы не допустить перекрестного опыления) и дал им возможность самоопылиться. Семена, собранные c растений F1, были пересчитаны и высажены следующей весной для получения второго гибридного поколения, F2 (поколение F2 - это всегда результат инбридинга в поколении F1, в данном случае самоопыления). Во втором гибридном поколении у одних растений образовались пазушные цветки, а у других - верхушечные. Иными словами, признак «верхушечные цветки», отсутствовавший в поколении F1, вновь появился в поколении F2. Мендель рассудил, что этот признак присутствовал в поколении F1 в скрытом виде, но не смог проявиться; поэтому он назвал его рецессивным. Из 858 растений, полученных Менделем в F2, у 651 были пазушные цветки, а у 207-верхушечные. Мендель провел ряд аналогичных опытов, используя всякий раз одну пару альтернативных признаков. Результаты экспериментальных скрещиваний по семи парам таких признаков приведены в табл. 1.
Таблица 1. Результаты экспериментов Менделя по наследованию семи пар альтернативных признаков. (Наблюдаемое соотношение доминантных и рецессивных признаков приближается к теоретически ожидаемому 3 : 1). Во всех случаях анализ результатов показал, что отношение доминантных признаков к рецессивным в поколении F2 составляло примерно 3 : 1. Приведенный выше пример типичен для всех экспериментов Менделя, в которых изучалось наследование одного признака (моногибридные скрещивания). На основании этих и аналогичных результатов Мендель сделал следующие выводы:
Теперь мы знаем, что эти факторы, детерминирующие такие признаки, как расположение цветка, соответствуют участкам хромосомы, называемым генами. Описанные выше эксперименты, проводившиеся Менделем при изучении наследования одной пары альтернативных признаков, служат примером моногибридного скрещивания.
Возвратное, или анализирующее, скрещиваниеОрганизм из поколения F1, полученного от скрещивания между гомозиготной доминантной и гомозиготной рецессивной особями, гетерозиготен по своему генотипу, но обладает доминантным фенотипом. Для того чтобы проявился рецессивный фенотип, организм должен быть гомозиготным по рецессивному аллелю. В поколении F2 особи с доминантным фенотипом могут быть как гомозиготами, таки гетерозиготами. Если селекционеру понадобилось выяснить генотип такой особи, то единственным способом, позволяющим сделать это, служит эксперимент с использованием метода, называемого анализирующим ( возвратным ) скрещиванием. Скрещивая организм неизвестного генотипа с организмом, гомозиготным по рецессивному аллелю изучаемого гена, можно определить этот генотип путем одного скрещивания. Например, у плодовой мушки Drosophila длинные крылья доминируют над зачаточными. Особь с длинными крыльями может быть гомозиготной (LL) или гетерозиготной (Ll). Для установления ее генотипа надо провести анализирующее скрещивание между этой мухой и мухой, гомозиготной по рецессивному аллелю (ll). Если у всех потомков от этого скрещивания будут длинные крылья, то особь с неизвестным генотипом - гомозигота по доминантному аллелю. Численное соотношение потомков с длинными и с зачаточными крыльями 1 : 1 указывает на гетерозиготность особи с неизвестным генотипом.
Дигибридное скрещивание и закон независимого распределенияУстановив возможность предсказывать результаты скрещиваний по одной паре альтернативных признаков, Мендель перешел к изучению наследования двух пар таких признаков. Скрещивания между особями, различающимися по двум признакам, называют дигибридными. В одном из своих экспериментов Мендель использовал растения гороха, различающиеся по форме и окраске семян. Применяя метод, описанный в разд. 2.1, он скрещивал между собой чистосортные ( гомозиготные) растения с гладкими желтыми семенами и чистосортные растения с морщинистыми зелеными семенами. У всех растений F1 (первого поколения гибридов) семена были гладкие и желтые. По результатам проведенных ранее моногибридных скрещиваний Мендель уже знал, что эти признаки доминантны; теперь, однако, его интересовали характер и соотношение семян разных талов в поколении F2, полученном от растений F1 путем самоопыления. Всего он собрал от растений F2 556 семян, среди которых было
гладких желтых 315
Соотношение разных фенотипов составляло примерно 9: 3: 3: 1 (дигибридное расщепление). На основании этих результатов Мендель сделал два вывода:
Краткое изложение сути гипотез Менделя
Хромосомная теория наследственностиК концу XIX в. в результате повышения оптических качеств микроскопов и совершенствования цитологических методов возможно стало наблюдать поведение хромосом в гаметах и зиготах. Еще в 1875 г. Гертвиг обратил внимание на то, что при оплодотворении яиц морского ежа происходит слияние (двух ядер - ядра спермия и ядра яйцеклетки. В 1902 г. Бовери продемонстрировал важную роль ядра в (регуляции развития признаков организма, а в 1882 г. Флемминг описал поведение хромосом во время митоза. В 1900 г. законы Менделя были вторично открыты и должным образом оценены почти одновременно и независимо друг от друга тремя учеными - де Фризом, Корренсом и Чермаком. Корренс сформулировал выводы Менделя в привычной нам форме двух законов и ввел термин «фактор», тогда как Мендель для описания единицы наследственности пользовался словом «элемент». Позднее американец Уильям Сэттон заметил удивительное сходство между поведением хромосом во время образования гамет и оплодотворения и передачей менделевских наследственных факторов. На основании изложенных выше данных Сэттон и Бовери высказали мнение, что хромосомы являются носителями менделевских факторов, и сформулировали так называемую хромосомную теорию наследственности. Согласно этой теории, каждая пара факторов локализована в паре гомологичных хромосом, причем каждая хромосома несет по одному фактору. Поскольку число признаков у любого организма во много раз больше числа его хромосом, видимых в микроскоп, каждая хромосома должна содержать множество факторов. В 1909 г. Иогансен заменил термин фактор, означавший основную единицу наследственности, термином ген. Альтернативные формы гена, определяющие его проявление в фенотипе, назвали аллеля- ми. Аллели - это конкретные формы, которыми может быть представлен ген, и они занимают одно и то же место - локус - в гомологичных хромосомах.
СцеплениеВсе ситуации и примеры, обсуждавшиеся до сих пор, относились к наследованию генов, находящихся в разных хромосомах. Как выяснили цитологи, у человека все соматические клетки содержат по 46 хромосом. Поскольку человек обладает тысячами различных признаков - таких, например, как группа крови, цвет глаз, способность секретировать инсулин, - в каждой хромосоме должно находиться большое число генов. Гены, лежащие в одной и той же хромосоме, называют сцепленными. Все гены какой-либо одной хромосомы образуют группу сцепления; они обычно попадают в одну гамету и наследуются вместе. Таким образом, гены, принадлежащие к одной группе сцепления, обычно не подчиняются менделевскому принципу независимого распределения. Поэтому при дигибридном скрещивании они не дают ожидаемого отношения 9:3:3:1. В таких случаях получаются самые разнообразные соотношения. У дрозофилы гены, контролирующие окраску тела и длину крыла, представлены следующими парами аллелей (назовем соответствующие признаки): серое тело - черное тело, длинные крылья - зачаточные (короткие) крылья. Серое тело и длинные крылья доминируют. Ожидаемое отношение фенотипов в F2 от скрещивания между гомозиготой с серым телом и длинными крыльями и гомозиготой с черным телом и зачаточными крыльями должно составить 9: 3: 3: 1. Это указывало бы на обычное менделевское наследование при дигибридном скрещивании, обусловленное случайным распределением генов, находящихся в разных, негомологичных хромосомах. Однако вместо этого в F2 были получены в основном родительские фенотипы в отношении примерно 3: 1. Это можно объяснить, предположив, что гены окраски тела и длины крыла локализованы в одной и той же хромосоме, т.е. сцеплены. Практически, однако, соотношение 3:1 никогда не наблюдается, а возникают все четыре фенотипа. Это объясняется тем, что колкое сцепление встречается редко. В большинстве экспериментов по скрещиванию при наличии сцепления помимо мух с родительскими фенотипами обнаруживаются особи с новыми сочетаниями признаков. Эти новые фенотипы называют рекомбинантными. Все это позволяет дать следующее определение сцепления: два или более генов называют сцепленными, если потомки с новыми генными комбинациями (рекомбинанты) встречаются реже, чем родительские фенотипы.
Группы сцепления и хромосомыГенетические исследования, проводившиеся в начале нашего века, в основном были направлены на выяснение роли генов в передаче признаков. Работы Моргана с плодовой мушкой Drosophila melanogaster показали, что большинство фенотипических признаков объединено у нее в четыре группы сцепления и признаки каждой группы наследуются совместно. Было замечено, что число групп сцепления соответствует числу пар хромосом. Изучение других организмов привело к сходным результатам. При экспериментальном скрещивании разнообразных организмов обнаружилось, что некоторые группы сцепления больше других (т.е. в них больше генов). Изучение хромосом этих организмов показало, что они имеют разную длину. Морган доказал наличие четкой связи между этими наблюдениями. Они послужили дополнительными подтверждениями локализации генов в хромосомах.
Гигантские хромосомы и геныВ 1913 г. Стертевант начал свою работу по картированию положения генов в хромосомах дрозофилы, во это было за 21 год до того, как появилась возможность связать различимые в хромосомах структуры с генами. В 1934 г. было замечено, что в клетках слюнных желез дрозофилы хромосомы примерно в 100 раз крупнее, чем в других соматических клетках. По каким-то причинам эти хромосомы многократно удваиваются, но не отделяются друг от друга, до тех пор пока их не наберется несколько тысяч, лежащих бок о бок. Окрасив хромосомы и изучая их с помощью светового микроскопа, можно увидеть, что они состоят из чередующихся светлых и темных поперечных полос. Для каждой хромосомы характерен свой особый рисунок полос. Первоначально предполагали, что эти полосы представляют собой гены, но оказалось, что дело обстоит не так просто. У дрозофилы можно искусственным путем вызывать различные фенотипические аномалии, которые сопровождаются определенными изменениями в рисунке поперечных полос, видимых под микроскопом. Эти фенотипические и хромосомные аномалии коррелируют в свою очередь с генными локусами. Это позволяет сделать вывод, что полосы на хромосомах действительно как-то связаны с генами, но взаимоотношения между теми и другими остаются пока неясными.
Определение пола
Рисунок 1. Хромосомные наборы самца и самки D. melanogaster. Они состоят из четырех пар хромосом (пара I - половые хромосомы).
Рисунок 2. Вид половых хромосом человека в метафазе митоза. Особенно четким примером описанного выше метода установления зависимости между фенотипи- ческими признаками организмов и строением их хромосом служит определение пола. У дрозофилы фенотипические различия между двумя полами явно связаны с различиями в хромосомах (рис. 1). При изучении хромосом у самцов и самок ряда животных между ними были обнаружены некоторые различия. Как у мужских, так и у женских особей во всех клетках имеются пары одинаковых (гомологичных) хромосом, но по одной паре хромосом они различаются. Это валовые хромосомы (гетеросомы). Все остальные хромосомы называют аутосомами. Как можно видеть на рис. 1, у дрозофилы четыре пары хромосом. Три пары (II, III и IV) идентичны у обоих полов, но пара I, состоящая из идентичных хромосом у самки, различается у самца. Эти хромосомы называют X - и Y - хромосомами; генотип самки XX, а самца - XY. Такие различия по половым хромосомам характерны для большинства животных, в том числе для человека (рис. 1), но у птиц (включая кур) и у бабочек наблюдается обратная картина: у самок имеются хромосомы XY, а у самцов - XX. У некоторых насекомых, например у прямокрылых, Y - хромосомы нет вовсе, так что самец имеет генотип ХО. При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. Например, у млекопитающих каждое яйцо содержит одну Х - хромосому, половина спермиев - одну X - хромосому, а другая половина - одну Y - хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. Пол с генотипом XX называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х - хромосомы, а пол с генотипом XY - гетерогаметным, так как половина гамет содержит X-, а половина - Y - хромосому. У человека генотипический пол данного индивидуума определяют, изучая неделящиеся клетки. Одна Х - хромосома всегда оказывается в активном состоянии и имеет обычный вид. Другая, если она имеется, бывает в покоящемся состоянии, в виде плотного темно - окрашенного тельца, называемого тельцем Барра. Число телец Барра всегда на единицу меньше числа наличных Х - хромосом, т.е. у самца (XY) их нет вовсе, а у самки (ХХ) - только одно. Функция Y - хромосомы, очевидно, варьирует в зависимости от вида. У человека Y - хромосома контролирует дифференцировку семенников, которая в дальнейшем влияет на развитие половых органов и мужских признаков. У большинства организмов, однако, Y - хромосома не содержит генов, имеющих отношение к полу. Ее даже называют генетически инертной или генетически пустой, так как в ней очень мало генов. Как полагают, у дрозофилы гены, определяющие мужские признаки, находятся в аутосомах, и их фенотипические эффекты маскируются наличием пары Х - хромосом; в присутствии одной Х - хромосомы мужские признаки проявляются. Это пример наследования, ограниченного полом (в отличие от наследования, сцепленного с полом), при котором, например, у женщин подавляются гены, детерминирующие рост бороды. Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в F1, получали равное число красноглазых самок и белоглазых самцов. Однако при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки. При скрещивании этих мух F1, между собой были получены красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой самки. Тот факт, что у самцов частота проявления рецессивного признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х - хромосоме, а Y - хромосома лишена гена окраски глаз. Чтобы проверить эту гипотезу, Морган скрестил исходного белоглазого самца с красноглазой самкой из F1. В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган справедливо заключил, что только Х - хромосома несет ген окраски глаз. В Y - хромосоме соответствующего локуса вообще нет. Это явление известно под названием наследования, сцепленного с полом.
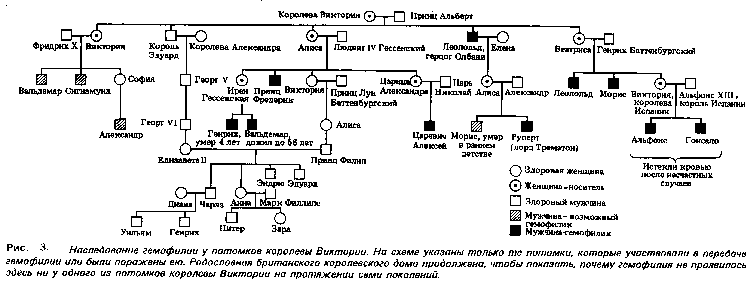
Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х - хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом, например цветовой слепоты, раннего облысения и гемофилии у человека. Гемофилия - сцепленный с полом рецессивный признак, при котором нарушается образование фактора VIII, ускоряющего свертывание крови. Ген, детерминирующий синтез фактора VIII, находится в участке Х - хромосомы, не имеющем гомолога, и представлен двумя аллелями - доминантным нормальным и рецессивным мутантным.
Возможны следующие генотипы и фенотипы:
Один из наиболее хорошо документированных примеров наследования гемофилии мы находим в родословной потомков английской королевы Виктории. Предполагают, что ген гемофилии возник в результате мутации у самой королевы Виктории или у одного из ее родителей. На рис. 3 показано, как этот ген передавался ее потомкам.
До сих пор рассматривались относительно простые аспекты генетики: доминирование, моногибридное и дигибридное скрещивание, сцепление, определение пола и наследование, сцепленное с полом. Известны, однако, и другие взаимодействия между генами, и возможно, что именно они определяют большую часть фенотипических признаков организма.
Известны случаи, когда два или более аллелей не проявляют в полной мере доминантность или рецессивность, так что в гетерозиготном состоянии ни один из аллелей не доминирует над другим. Это явление неполового доминирования, или кодоминантность, представляет собой исключение из описанного Менделем правила наследования при моногибридных скрещиваниях. К счастью, Мендель выбрал для своих экспериментов признаки, которым не свойственно неполное доминирование; в противном случае оно могло бы сильно осложнить его первые исследования.
Неполное доминирование наблюдается как у растений, так и у животных. В большинстве случаев гетерозиготы обладают фенотипом, промежуточным между фенотипами доминантной и рецессивной гомозигот. Примером служат андалузские куры, полученные в результате скрещивания чистопородных черных и «обрызганных белых» (splashed white) кур. Черное оперение обусловлено наличием аллеля, определяющего синтез черного пигмента меланина. У «обрызганных» кур этот аллель отсутствует. У гетерозигот меланин развивается не в полной мере, создавая лишь голубоватый отлив на оперении.
Поскольку общепринятых символов для обозначения аллелей с неполным доминированием не существует, нам необходимо ввести для генотипов такие символы, чтобы сделать понятными приведенные ниже схемы получения андалузских кур.
Возможны, например, такие обозначения: черные - В, «обрызганные» - b, W, BW или BBW. Результаты скрещивания между гомозиготными черными и «обрызганными» курами представлены в табл. 2.
При скрещивании между собой особей F1 отношение фенотипов в F2 отличается от менделевского отношения 3 : 1, типичного для моногибридного скрещивания. В этом случае получается отношение 1:2:1, где у половины особей F2 будет такой же генотип, как у F1 (табл. 3). Отношение 1 : 2 : 1 характерно для результатов скрещиваний при неполном доминировании.
Таблица 2. Скрещивание кур андалузской породы: гибриды F1.
Известны случаи, когда один ген может оказывать влияние на несколько признаков, в том числе на жизнеспособность. У человека и других млекопитающих определенный рецессивный ген вызывает образование внутренних спаек легких, что приводит к смерти при рождении. Другим примером служит ген, который влияет на формирование хряща и вызывает врожденные уродства, ведущие к смерти плода или новорожденного.
У кур, гомозиготных по аллелю, вызывающему «курчавость» перьев, неполное развитие перьев влечет за собой несколько фенотипических эффектов. У таких кур теплоизоляция недостаточна, и они страдают от охлаждения. Для компенсации потери тепла у них появляется ряд структурных и физиоло-
гических адаптаций, но эти адаптации малоэффектны и среди таких кур высока смертность.
Воздействие летального гена ясно видно на примере наследования окраски шерсти у мышей. У диких мышей шерсть обычно серая, типа агути; но у некоторых мышей шерсть желтая. При скрещиваниях между желтыми мышами в потомстве получаются как желтые мыши, так и агути в отношении 2 : 1. Единственное возможное объяснение таких результатов состоит в том, что желтая окраска шерсти доминирует над агути и что все желтые мыши гетерозиготны. Атипичное менделевское отношение объясняется гибелью гомозиготных желтых мышей до рождения. При вскрытии беременных желтых мышей, скрещенных с желтыми же мышами, в их матках были обнаружены мертвые желтые мышата. Если же скрещивались желтые мыши и агути, то в матках беременных самок не оказывалось мертвых желтых мышат, поскольку при таком скрещивании не может быть потомства, гомозиготного по гену желтой шерсти.
Ген называют эпистатическим (от греч. еpi - над), если его присутствие подавляет эффект какого-либо гена, находящегося в другом локусе. Эпистатические гены иногда называют ингибирующими генами, а те гены, действие которых ими подавляется, - гипостатическими (от греч. hypo - под).
Окраска шерсти у мышей контролируется парой генов, находящихся в разных локусах. Эпистатический ген определяет наличие окраски и имеет два аллеля: доминантный, определяющий окрашенную шерсть, и рецессивный, обусловливающий альбинизм (белая окраска). Гипостатический ген определяет характер окраски и имеет два аллеля: агути (доминантный, определяющий серую окраску) и черный (рецессивный). Мыши могут иметь серую или черную окраску в зависимости от своих генотипов, но наличие окраски возможно только в том случае, если у них одновременно имеется аллель окрашенной шерсти. Мыши, гомозиготные по рецессивному аллелю альбинизма, будут альбиносами даже при наличии у них аллелей агути и черной шерсти. Возможны три разных фенотипа: агути, черная шерсть и альбинизм. При скрещивании можно получить эти фенотипы в различных соотношениях в зависимости от генотипов скрещиваемых особей.
Многие из самых заметных признаков организма представляют собой результат совместного действия многих различных генов; эти гены образуют особый генный комплекс, называемый полигенной системой. Хотя вклад каждого отдельного гена, входящего в такую систему, слишком мал, чтобы оказать сколько-нибудь значительное влияние на фенотип, почти бесконечное разнообразие, создаваемое совместным действием этих генов (полигенов), составляет генетическую основу непрерывной изменчивости.
Изменчивостью называют всю совокупность различий по тому или иному признаку между организмами, принадлежащими к одной и той же природной популяции или виду. Поразительное морфологическое разнообразие особей в пределах любого вида привлекло внимание Дарвина и Уоллеса во время их путешествий. Закономерный, предсказуемый характер передачи таких различий по наследству послужил основой для исследований Менделя. Дарвин установил, что определенные признаки могут развиваться в результате отбора, тогда как Мендель объяснил механизм, обеспечивающий передачу из поколения в поколение признаков, по которым ведется отбор.
Мендель описал, каким образом наследственные факторы определяют генотип организма, который в процессе развития проявляется в структурных, физиологических и биохимических особенностях фенотипа. Если фенотипическое проявление любого признака обусловлено в конечном счете генами, контролирующими этот признак, то на степень развития определенных признаков может оказывать влияние среда.
Изучение фенотипических различий в любой большой популяции показывает, что существуют две формы изменчивости - дискретная и непрерывная. Для изучения изменчивости какого-либо признака, например роста у человека, необходимо измерить этот признак у большого числа индивидуумов в изучаемой популяции. Результаты измерений представляют в виде гистограммы, отражающей распределение частот различных вариантов этого признака в популяции. На рис. 4 представлены типичные результаты, получаемые при таких исследованиях, и они наглядно демонстрируют различие между дискретной и непрерывной изменчивостью.
Некоторые признаки в популяции представлены ограниченным числом вариантов. В этих случаях различия между особями четко выражены, а промежуточные формы отсутствуют; к таким признакам относятся, например, группы крови у человека, длина крыльев у дрозофилы, меланистическая и светлая формы у березовой пяденицы (Biston betularia), длина столбика у первоцвета (Primula) и пол у животных и растений. Признаки, для которых характерна дискретная изменчивость, обычно контролируются одним или двумя главными генами, у которых может быть два или несколько аллелей, и внешние условия относительно мало влияют на их фенотипическую экспрессию.
Поскольку дискретная изменчивость ограничена некоторыми четко выраженными признаками, ее называют также качественной изменчивостью в отличие от количественной, или непрерывной, изменчивости.
Рисунок 4. Гистограммы, отражающие распределение частот в случае прерывистой (А) и не прерывистой (Б) изменчивости.
По многим признакам в популяции наблюдается полный ряд переходов от одной крайности к другой без всяких разрывов. Наиболее яркими примерзлая служат такие признаки, как масса (вес), линейные размеры, форма и окраска организма в целом или отдельных его частей. Частотное распределение по признаку, проявляющему непрерывную изменчивость, соответствует кривой нормального распределения. Большинство членов популяции попадает в среднюю часть кривой, а на ее концах, соответствующих двум крайним значениям данного признака, находится примерное одинаковое (очень малое) число особей. Признаки, для которых характерна непрерывная изменчивость, обусловлены совместным воздействием многих генов (полигенов) и факторов среды. Каждый из этих генов в отдельности оказывает очень небольшое влияние на фенотип, но совместно они создают значительный эффект.
Главный фактор, детерминирующий любой фенотипический признак, - это генотип. Генотип организма определяется в момент оплодотворения, но степень последующей экспрессии этого генетического потенциала в значительной мере зависит от внешних факторов, воздействующих на организм во время его развития. Так, например, использованный Менделем сорт гороха с длинным стеблем обычно достигал высоты 180 см. Однако для этого ему необходимы были соответствующие условия - освещение, снабжение водой и хорошая почва. При отсутствии оптимальных условий (при наличии лимитирующих факторов) ген высокого стебля не мог в полной мере проявить свое действие. Эффект взаимодействия генотипа и факторов среды продемонстрировал датский генетик Иогансен. В ряде экспериментов на карликовой фасоли он выбирал из каждого поколения самоопылявшихся растений самые тяжелые и самые легкие семена и высаживал их для получения следующего поколения. Повторяя эти эксперименты на протяжении нескольких лет, он обнаружил, что в пределах «тяжелой» или «легкой» селекционной линии семена мало различались по среднему весу, тогда как средний вес семян из разных линий сильно различался. Это позволяет считать, что на фенотипическое проявление признака оказывают влияние как наследственность, так и среда. На основании этих результатов можно определить непрерывную фенотипическую изменчивость как «кумулятивный эффект варьирующих факторов среды, воздействующих на вариабельный генотип». Кроме того, эти результаты показывают, что степень наследуемости данного признака определяется в первую очередь генотипом. Что касается развития таких чисто человеческих качеств, как индивидуальность, темперамент и интеллект, то, судя по имеющимся данным, они зависят как от наследственных, так и от средовых факторов, которые, взаимодействуя в различной степени у разных индивидуумов, влияют на окончательное выражение признака. Именно эти различия в тех и других факторах создают фенотипические различия между индивидуумами. Мы пока еще не располагаем данными, которые твердо указывали бы на то, что влияние каких-то из этих факторов всегда преобладает, однако среда никогда не может вывести фенотип за пределы, детерминированные генотипом.
Необходимо ясно представлять себе, что взаимодействие между дискретной и непрерывной изменчивостью и средой делает возможным существование двух организмов с идентичным фенотипом. Механизм репликации ДНК при митозе столь близок к совершенству, что возможности генетической изменчивости у организмов с бесполым размножением очень малы. Поэтому любая видимая изменчивость у таких организмов почти наверное обусловлена воздействиями внешней среды. Что же касается организмов, размножающихся половым путем, то у них есть широкие возможности для возникновения генетических различий. Практически неограниченными источниками генетической изменчивости служат два процесса, происходящие во время мейоза:
1. Реципрокный обмен генами между хромата- дамп гомологичных хромосом, который может происходить в профазе 1 мейоза. Он создает новые группы сцепления, т.е. служит важным источником генетической рекомбинации аллелей.
2. Ориентация пар гомологичных хромосом (бивалентов) в экваториальной плоскости веретена в метафазе I мейоза определяет направление, в котором каждый член пары будет перемещаться в анафазе I. Эта ориентация носит случайный характер. Во время метафазы II пары хроматид опять- таки ориентируется случайным образом, и этим определяется, к какому из двух противоположных полюсов направится та или иная хромосома во время анафазы II. Случайная ориентация и последующее независимое расхождение (сегрегация) хромосом делают возможным большое число различных хромосомных комбинаций в гаметах; число это можно подсчитать.
Третий источник изменчивости при половом размножении - это то, что слияние мужских и женских гамет, приводящее к объединению двух гаплоидных наборов хромосом в диплоидном ядре зиготы, происходит совершенно случайным образом (во всяком случае, в теории); любая мужская гамета потенциально способна слиться с любой женской гаметой.
Эти три источника генетической изменчивости и обеспечивают постоянную «перетасовку» генов, лежащую в основе происходящих все время генетических изменений. Среда оказывает воздействие на весь ряд получающихся таким образом фенотипов, и те из них, которые лучше всего приспособлены к данной среде, преуспевают. Это ведет к изменениям частот аллелей и генотипов в популяции. Однако эти источники изменчивости не порождают крупных изменений в генотипе, которые необходимы, согласно эволюционной теории, для возникновения новых видов. Такие изменения возникают в результате мутаций.
Мутацией называют изменение количества или структуры ДНК данного организма. Мутация приводит к изменению генотипа, которое может быть унаследовано клетками, происходящими от мутант- ной клетки в результате митоза или мейоза. Мутирование может вызывать изменения каких-либо признаков в популяции. Мутации, возникшие в половых клетках, передаются следующим поколениям организмов, тогда как мутации в соматических клетках наследуются только дочерними клетками, образовавшимися путем митоза, и такие мутации называют соматическими.
Мутации, возникающие в результате изменения числа или макроструктуры хромосом, известны под названием хромосомных мутаций или хромосомных аберраций (перестроек). Иногда хромосомы так сильно изменяются, что это можно увидеть под микроскопом. Но термин «мутация» используют главным образом для обозначения изменения структуры ДНК в одном докую, когда происходит так называемая генная, или точечная, мутация.
Представление о мутации как о причине внезапного появления нового признака было впервые выдвинуто в 1901 г. голландским ботаником Гуго де Фризом, изучавшим наследственность у энотеры Oenothera lamarckiana. Спустя 9 лет Т. Морган начал изучать мутации у дрозофилы, и вскоре при участии генетиков всего мира у нее было идентифицировано более 500 мутаций.
Внезапные спонтанные изменения фенотипа, которые нельзя связать с обычными генетическими явлениями или микроскопическими данными о наличии хромосомных аберраций, можно объяснить только изменениями в структуре отдельных генов. Генная, или точечная (поскольку она относится к определенному генному локусу), мутация - результат изменения нуклеотидной последовательности молекулы ДНК в определенном участке хромосомы. Такое изменение последовательности оснований в данном гене воспроизводится при транскрипции в структуре мРНК и приводит к изменению последовательности аминокислот в полипептидной цепи, образующейся в результате трансляции на рибосомах.
Существуют различные типы генных мутаций, связанных с добавлением, выпадением или перестановкой оснований в гене. Это дупликации, вставки, делении, инверсии или замены оснований. Во всех случаях они приводят к изменению нуклеотидной последовательности, а часто - и к образованию измененного полипептида. Например, делеция вызывает сдвиг рамки.
Генные мутации, возникающие в гаметах или в будущих половых клетках, передаются всем клеткам потомков и могут влиять на дальнейшую судьбу популяции. Соматические генные мутации, происходящие в организме, наследуются только теми клетками, которые образуются из мутантной клетки путем митоза. Они могут оказать воздействие на тот организм, в котором они возникли, но со смертью особи исчезают из генофонда популяции. Соматические мутации, вероятно, возникают очень часто и остаются незамеченными, но в некоторых случаях при этом образуются клетки с повышенной скоростью роста и деления. Эти клетки могут дать начало опухолям - либо доброкачественным, которые не оказывают особого влияния на весь организм, либо злокачественным, что приводит к раковым заболеваниям.
Эффекты генных мутаций чрезвычайно разнообразны. Большая часть мелких генных мутаций фенотипически не проявляется, поскольку они рецессивны, однако известен ряд случаев, когда изменение всего лишь одного основания в определенном гене оказывает глубокое влияние на фенотип. Одним из примеров служит серповидноклеточная анемия - заболевание, вызываемое у человека заменой основания в одном из генов, ответственных за синтез гемоглобина. Молекула дыхательного пигмента гемоглобина у взрослого человека состоит из четырех полипептидных цепей (двух α- и двух ß- цепей), к которым присоединены четыре простетические группы гема. От структуры полипептидных цепей зависит способность молекулы гемоглобина переносить кислород. Изменение последовательности оснований в триплете, кодирующем одну определенную аминокислоту из 146, входящих в состав ß-цепей, приводит к синтезу аномального гемоглобина серповидных клеток (HbS). Последовательности аминокислот в нормальных и аномальных ß-цепях различаются тем, что в одной точке аномальных цепей гемоглобина S глутамидовая кислота замещена валином. В результате такого, казалось бы, незначительного изменения гемоглобин S кристаллизуется при низких концентрациях кислорода, а это в свою очередь приводит к тому, что в венозной крови эритроциты с таким гемоглобином деформируются (из округлых становятся серповидными) и быстро разрушаются. Физиологический эффект мутации состоит в развитии острой анемии и снижении количества кислорода, переносимого кровью. Анемия не только вызывает физическую слабость, но и может привести к нарушениям деятельности сердца и почек и к ранней смерти людей, гомозиготных по мутантному аллелю. В гетерозиготном состоянии этот аллель вызывает значительно меньший эффект: эритроциты выглядят нормальными, а аномальный гемоглобин составляет только около 40%. У гетерозигот развивается анемия лишь в слабой форме, а зато в тех областях, где широко распространена малярия, особенно в Африке и Азии, носители аллеля серповидноклеточности невосприимчивы к этой болезни. Это объясняется тем, что ее возбудитель -малярийный плазмодий - не может жить в эритроцитах, содержащих аномальный гемоглобин.
Хромосомные и генные мутации оказывают разнообразные воздействия на организм. Во многих случаях эти мутации летальны, так как нарушают развитие; у человека, например, около 20% беременностей заканчиваются естественным выкидышем в сроки до 12 недель, и в половине таких случаев можно обнаружить хромосомные аномалии. В результате некоторых хромосомных мутаций определенные гены могут оказаться вместе, и их общий эффект может привести к появлению какого-либо «благоприятного» признака. Кроме того, сближение некоторых генов друг с другом делает менее вероятным их разделение в результате кроссинговера, а в случае благоприятных генов это создает преимущество.
Генная мутация может привести к тому, что в определенном локусе окажется несколько аллелей. Это увеличивает как гетерозиготность данной популяции, так и ее генофонд, и ведет к усилению внутрипопуляционной изменчивости. Перетасовка генов как результат кроссинговера, независимого распределения, случайного оплодотворения и мутаций может повысить непрерывную изменчивость, но ее эволюционная роль часто оказывается преходящей, так как возникающие при этом изменения могут быстро сгладиться вследствие «усреднения». Что же касается генных мутаций, то некоторые из них увеличивают дискретную изменчивость, и это может оказать на популяцию более глубокое влияние. Большинство генных мутаций рецессивны по отношению к «нормальному» аллелю, который, успешно выдержав отбор на протяжении многих поколений, достиг генетического равновесия с остальным генотипом. Будучи рецессивными, мутантные аллели могут оставаться в популяции в течение многих поколений, пока им не удастся встретиться, т. е. оказаться в гомозиготном состоянии и проявиться в фенотипе. Время от времени могут возникать и доминантные мутантные оплели, которые немедленно дают фенотипический эффект.
Роль генов в развитии организма огромна. Гены характеризуют все признаки будущего организма, такие, как цвет глаз и кожи, размеры, вес и многое другое. Гены являются носителями наследственной информации, на основе которой развивается организм.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||